蔗糖与谷氨酰胺对巴尔通体液体培养影响的再评估
2 结果
2.1 谷氨酰胺添加对照实验
在基础液体培养基中添加2 mmol/L谷氨酰胺,通过Biosense系统实时监测汉赛巴尔通体、五日热巴尔通体两种不同宿主巴尔通体的生长情况。从生长曲线看,添加谷氨酰胺的汉赛巴尔通体在对数生长期和稳定期的OD值均高于未添加的(图1),但没有显著性差异(P > 0.05);五日热巴尔通体在添加谷氨酰胺后细菌量在对数生长期和稳定期略微有所增加,但影响不大(P > 0.05)。
2.2 蔗糖添加对照实验
选用汉赛巴尔通体和五日热巴尔通体观察液体培养基中添加蔗糖后细菌生长曲线的变化。结果发现,添加蔗糖对这两种巴尔通体生长有一定影响,但并不显著,P > 0.05(表2、3)。
从生长曲线看,汉赛巴尔通体和五日热巴尔通体生长速度不同,对添加蔗糖的反应也不同(图2A、2B、2C、2D)。汉赛巴尔通体对蔗糖不敏感,培养液中添加蔗糖后生长基本趋势没有变化(图1A、1C)。添加蔗糖使五日热巴尔通体在对数生长期浓度略高,稳定期后菌液浓度低于未添加蔗糖的培养液(图1B、1D)。
相比汉赛巴尔通体在接种后约12 h进入对数生长期(Biosense系统显示),五日热巴尔通体几乎没有停滞期直接进入对数生长期;在添加蔗糖的培养基中,五日热巴尔通体和汉赛巴尔通体分别在第3天和第4天间维持在稳定期;在未加蔗糖的培养基中,五日热巴尔通体接种后第4天菌液浓度达到最高,随即进入衰亡期。
培养第4天时测定每瓶培养液内全部细菌的湿重,添加与不添加蔗糖培养菌量分别为:汉赛巴尔通体是65.7 mg和68.5 mg,五日热巴尔通体是124.8 mg和120.3 mg。五日热巴尔通体的菌量明显高于汉赛巴尔通体(P < 0.01),添加蔗糖对这两种菌的收获量均没有明显影响,P > 0.05。
依据公式 G = (t₂ − t₁) / [(log W₁ − log W₂) / lg 2] 计算两种巴尔通体的世代时间,选择对数期斜率最大的两点作为 t₁ 和 t₂。汉赛巴尔通体依据第1天和第2天菌液浓度(CFU/mL)计算世代时间为5.2 h;五日热巴尔通体依据第2天和第3天菌液浓度(CFU/mL)计算世代时间为4.3 h。
在培养过程中,培养液的pH变化与生长曲线基本是一致的,对数期时pH处于上升期,从最初的5.5逐渐上升至8.0不再变化,添加蔗糖与否不影响这一变化过程。
表2 添加蔗糖对汉赛巴尔通体生长曲线的影响
| Culture days | x̄ ± sa (CFU/mL, OD600 values) | |
|---|---|---|
| +Sucb | −Succ | |
| 0 | 2.10 × 106 ± 2.65 × 105 (0.20 ± 0.00) | 2.10 × 106 ± 2.65 × 105 (0.20 ± 0.00) |
| 1 | 4.03 × 106 ± 5.77 × 105 (0.20 ± 0.00) | 3.27 × 106 ± 1.53 × 105 (0.20 ± 0.00) |
| 2 | 1.07 × 108 ± 2.52 × 107 (0.30 ± 0.06) | 8.00 × 107 ± 1.00 × 106 (0.26 ± 0.00) |
| 3 | 4.00 × 108 ± 1.00 × 108 (0.60 ± 0.00) | 6.00 × 108 ± 2.65 × 107 (0.80 ± 0.00) |
| 4 | 6.00 × 108 ± 1.00 × 108 (1.00 ± 0.06) | 7.33 × 108 ± 5.77 × 107 (1.16 ± 0.00) |
| 5 | 3.30 × 108 ± 1.53 × 108 (1.30 ± 0.06) | 2.13 × 108 ± 1.53 × 107 (1.44 ± 0.00) |
| 6 | 2.10 × 107 ± 1.00 × 106 (1.37 ± 0.06) | 2.60 × 107 ± 1.00 × 107 (1.51 ± 0.06) |
| 7 | 9.00 × 106 ± 3.61 × 106 (1.38 ± 0.06) | 4.30 × 106 ± 1.53 × 106 (1.51 ± 0.06) |
| 8 | 3.00 × 106 ± 0.00 (1.38 ± 0.06) | 1.70 × 106 ± 5.77 × 105 (1.51 ± 0.06) |
| 9 | 5.30 × 105 ± 1.15 × 105 (1.36 ± 0.06) | 4.70 × 105 ± 2.52 × 105 (1.46 ± 0.06) |
表3 添加蔗糖对五日热巴尔通体生长曲线的影响
| Culture days | x̄ ± sa (CFU/mL, OD600 values) | |
|---|---|---|
| +Sucb | −Succ | |
| 0 | 5.30 × 105 ± 5.77 × 104 (0.20 ± 0.00) | 5.30 × 105 ± 5.77 × 104 (0.20 ± 0.00) |
| 1 | 1.10 × 107 ± 1.41 × 106 (0.20 ± 0.00) | 3.00 × 106 ± 1.00 × 106 (0.20 ± 0.00) |
| 2 | 6.00 × 108 ± 1.73 × 108 (0.30 ± 0.00) | 4.30 × 107 ± 1.53 × 107 (0.20 ± 0.00) |
| 3 | 8.70 × 109 ± 1.53 × 109 (1.43 ± 0.06) | 2.00 × 109 ± 0.00 (0.47 ± 0.06) |
| 4 | 8.30 × 109 ± 2.31 × 109 (1.80 ± 0.00) | 1.57 × 1010 ± 5.03 × 109 (2.13 ± 0.12) |
| 5 | 1.47 × 109 ± 4.16 × 108 (1.70 ± 0.00) | 7.00 × 109 ± 2.08 × 108 (2.07 ± 0.06) |
| 6 | 8.70 × 107 ± 2.08 × 107 (1.70 ± 0.00) | 5.30 × 108 ± 5.77 × 107 (2.07 ± 0.15) |
| 7 | 8.00 × 106 ± 2.83 × 106 (1.70 ± 0.00) | 1.57 × 108 ± 2.52 × 107 (2.03 ± 0.12) |
| 8 | 1.10 × 106 ± 4.00 × 105 (1.63 ± 0.06) | 2.20 × 107 ± 2.65 × 106 (2.00 ± 0.10) |
| 9 | 3.00 × 103 ± 5.77 × 102 (1.60 ± 0.00) | 5.30 × 105 ± 1.53 × 105 (1.93 ± 0.12) |
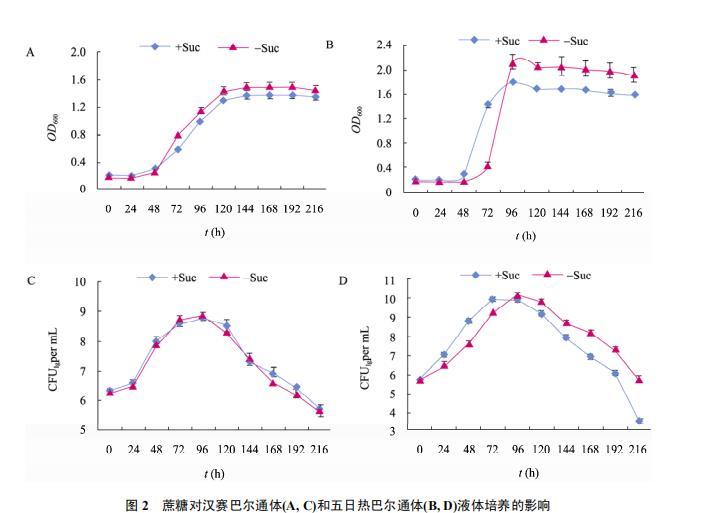
注: 起始接种浓度为 2.0 OD; 图 A、B 和 C、D 分别以每 24 h 细菌培养液 OD 值变化和生长量(CFU/mL)绘制生长曲线.
2.3 其他巴尔通体生长曲线的测定
由于Bc、Bk、Bb、Bvb、Bva、Bvv、Be、Bg、Bd和Bt在固体培养基上生长速度不同,为掌握这些巴尔通体在液体培养时的状态,本研究采用Biosense系统观察记录了它们在液体培养中的生长情况并绘制生长曲线。
从生长曲线看(图3),Bvb、Bva、Bvv、Be、Bg、Bd和Bt在培养液中生长良好,大部分菌株在培养0–1 d处于停滞期,1–4 d处于对数期;Be和Bva较其他菌生长更快;Bvv生长略慢,培养0–2 d处于停滞期,2–6 d处于对数期。
Bk在培养液中呈贴壁生长,在培养第3天即可见瓶壁上贴附的白色菌体,不易脱落,用力摇动瓶体或强行刮下,在培养液中形成团块或颗粒,不易混匀,导致Biosense系统读数波动较大,但仍可观察到生长趋势。Bc在液体培养基中未见生长。
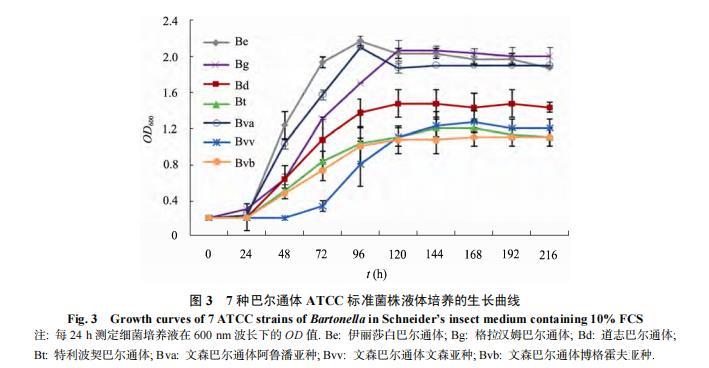
(图注:曲线由Biosense系统实时监测获得,每30 min记录一点。菌株包括:Be: 伊丽莎白巴尔通体;Bg: 格拉汉姆巴尔通体;Bd: 道志巴尔通体;Bt: 特利波契巴尔通体;Bva: 文森巴尔通体阿鲁潘亚种;Bvv: 文森巴尔通体文森亚种;Bvb: 文森巴尔通体博格霍夫亚种。)
3 讨论
由于巴尔通体对铁离子的依赖,在固体培养时需要添加血液成分(羊、兔、马或人全血),在已报道的数种液体培养方法中也需要添加类似成分,如动物全血、血红素或胎牛血清,还有一些其他成分如Fildes提取物、氨基酸和矿物盐等。
Riess等在研究巴尔通体与果蝇细胞相互作用时发现SIM添加FCS后适用于巴尔通体的培养,其他添加成分还有谷氨酰胺和蔗糖。谷氨酰胺是动物细胞培养时必需添加的氨基酸,巴尔通体可以将其分解为谷氨酸盐作为碳源和能量来源。本研究中应用的液体培养基是含L-谷氨酰胺SIM,那么在培养中是否有必要再添加此成分,我们对此做了进一步探讨。研究结果显示添加与否对巴尔通体生长无明显影响,因此,建议在应用这种含L-谷氨酰胺SIM时无需再次添加谷氨酰胺。关于蔗糖是否对巴尔通体生长有促进效果,本研究结果与Riess等的报道不一致,即添加蔗糖没有明显促进巴尔通体的生长。由于没有完全的糖酵解途径,巴尔通体不能利用葡萄糖作为能量来源,如何利用蔗糖及其是否能作为碳源或能量来源或其他作用,还需要进一步的研究,或许将能够解释结果不一致的原因。
在固体培养时汉赛巴尔通体和五日热巴尔通体的生长速度和菌量没有明显差别,而液体培养使五日热巴尔通体在这两方面都优于汉赛巴尔通体,菌量几乎是汉赛巴尔通体的2倍,通过计算世代时间,也可以看出五日热巴尔通体的繁殖速度快于汉赛巴尔通体,为什么液体培养会造成这种生长速度的差异,还有待于进一步研究。
巴尔通体生长的最适pH是6.8–7.2,从生长曲线看,这个范围是对数期,随着pH不断升高,巴尔通体的繁殖进入衰亡期。液体培养过程中pH的变化与分解氨基酸产生NH₃有关,氨基酸不断被消耗产生NH₃使培养液碱化,直至不再适于巴尔通体生长。因此,在传代培养时,应该在对数期或稳定期将菌体重新接入新的培养液中。另外,也可以将pH的变化作为指示,判断培养液中氨基酸的消耗和巴尔通体的生长状态。
本研究还采用Biosense系统对其他10种巴尔通体进行了液体培养的实时监测,发现鼠类宿主的巴尔通体较猫、犬和人作为宿主的巴尔通体生长速度要快,与固体培养是一致的。生长曲线反映出不同种巴尔通体具有不同繁殖速度,为进一步掌握这些巴尔通体的生长特性提供了高分辨率的基础数据。总的来说,在液体培养基中巴尔通体的生长比在固体培养基上快,特别是某些种,如克勒巴尔通体和杆菌样巴尔通体(数据未显示),在固体培养基上生长非常缓慢,通常需要10 d才能长出微小菌落,而在液体培养时生长明显加快。结合Biosense系统的实时监测能力,可大幅提高培养效率和数据精度,为这类更难培养的巴尔通体提供了很好的解决方案。
相关新闻推荐
1、李斯特菌ST87在25°C不同营养条件下牛肉和蔬菜汁中的生长曲线