罗尔阿太菌生长曲线、原生质体制备条件及融合技术(四)
2.4融合条件对原生质体融合率的影响
融合条件对原生质体融合率的影响见图3,由图3a可以看出,随着PEG6000浓度的增加,原生质体融合率先增加后降低。当PEG6000为0.4 g·mL——1时,原生质体融合率为6.792%.原因可能是PEG6000在原生质体融合过程中既作为诱导剂又作为稳定剂,浓度低时诱导作用不明显,浓度高时原生质体不稳定易破裂,致使原生质体融合率降低。
由图3b可以看出,随着融合时间的增加,原生质体融合率先增加后降低。当融合时间为25 min时,原生质体融合率为6.543%.可能原因是融合时间短,原生质体融合不完全。融合时间长,PEG6000会对细胞造成毒害作用,致使原生质体融合率降低。
由图3c可以看出,随着融合温度的增加,原生质体融合率先增加后降低。当融合温度为30℃时,原生质体融合率为6.452%.原因可能是融合温度低时达不到融合条件,融合温度高时抑制PEG6000作用,致使原生质体融合率降低。
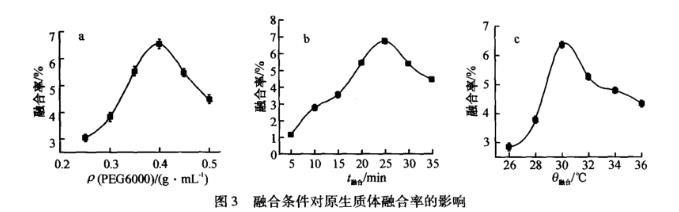
图3融合条件对原生质体融合率的影响
2.5融合子的筛选
2.5.1原生质体融合子形态
原生质体融合子形态见图4,原生质体经紫外和热处理灭活后,致死率为100%,在双层再生培养基上不再生长。原生质体融合后在再生培养基上长出融合子,最初是1个白色圆形的点,随后菌丝向外萌发生长,通常需要6——9 d.
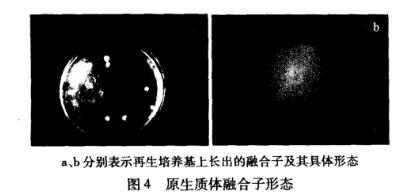
图4原生质体融合子形态
2.5.2筛选菌株多糖及草酸含量经原生质体融合筛选出了4个菌株,分别命名为AY-1、AY-2、AY-3、AY-4,在发酵培养基中培养5.5 d后测得多糖及草酸产量见表3.
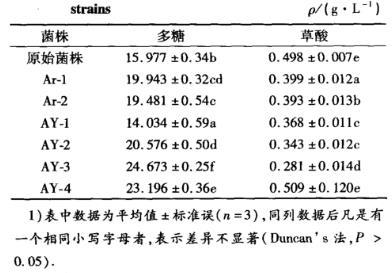
表3筛选菌株的多糖和草酸产量
表中数据为平均值±标准误(n=3),同列数据后凡是有一个相同小写字母者,表示差异不显著(Duncan’s法,P>0.05)。
由表3可以看出,经原生质体融合选育的菌株中,除菌株AY-1的多糖产量下降外,其他菌株多糖产量都高于Ar-1和Ar-2,菌株AY-3和AY-4多糖产量较高,分别比原始菌株的产糖量提高了54.42%和45.18%.此外,菌株AY-3草酸产量最低,为0.281 g·L——1,比原始菌株的草酸产量降低了43.57%.菌株AY-1、AY-2、AY-3、AY-4多糖产量均与原始菌株差异显著(P<0.05)。草酸产量除菌株AY-4与原始菌株差异不显著外(P>0.05),菌株AY-1、AY-2、AY-3均与原始菌株差异显著(P<0.05)。菌株AY-3的多糖产量、多糖产量变化、草酸产量、草酸产量变化均与其他菌株差异显著(P<0.05),为最佳产糖菌株。
3讨论与结论
真菌原生质体融合技术不仅打破了微生物种界界限,还能保持完整的遗传物质,从而完成菌株基因重组,重组后的融合子具有双亲遗传性状,能够表现出双亲的优良性状。本研究中以罗尔阿太菌菌株AY6657741为原始菌株,诱变株Ar-1和Ar-2为双亲菌株,以玉米淀粉和玉米黄浆为培养基,采用原生质体融合技术选育的罗尔阿太菌菌株多糖产量高达24.673 g·L——1.本试验中多糖产量与Shrikant等的研究相比,产糖量提高了10.54%.这表明原生质体融合技术能较好地提高罗尔阿太菌分泌多糖的能力,为今后进一步提高多糖产量奠定了基础。
本试验中采用紫外照射和热处理方法灭活双亲原生质体,但紫外处理和热处理在一定条件下有可逆性,因此融合培养后的菌株具有不稳定性,其性状特征可能很快失去,所以必须传代数次保证其性状的稳定性。另外在发酵过程中发现,伴随多糖的产生,会生成草酸,使发酵液pH由4.55很快降至2.00左右,从而抑制了多糖的分泌,当草酸产量高于0.368 g·L——1时,多糖产量就会受到影响。采用原生质体融合技术选育出的菌株AY-3满足了草酸低产,多糖高产的要求。
目前高产罗尔阿太菌多糖生产专利技术由Cargill公司拥有,我国只能依赖进口。本研究中获得的菌株AY-3可为我国获得拥有自主知识产权的罗尔阿太菌多糖工艺技术奠定了一定的理论基础。今后的研究将进一步探讨AY-3生产多糖的工艺,以期为该糖工业化生产奠定基础。
相关新闻推荐
1、不同温度下鲜切草鱼脊肉块中热杀索丝菌的生长规律、动力学模型的拟合及验证
2、沼泽红假单胞菌R1菌株的培养基配方、培养条件、生长量及影响因素(一)
3、生长曲线测定法和菌落计数法确定YchJ对鼠伤寒沙门菌抗逆能力的影响——摘要