草鱼呼肠孤病毒培养与滴度测定、及在草鱼、CIK细胞上的生长特性研究(三)
2.2 GCRV 873株在CIK细胞中的生长特性
GCRV 873株吸附CIK细胞1 h后,分别在0、2、4、8、10、12、24、36、48、72 h收取细胞病毒液。对各个时间点病毒滴度及RNA水平进行检测(图3),并通过显微镜观察不同时间点细胞病变情况(图4)。由图3可以看出病毒在感染CIK细胞0~12 h期间,病毒RNA水平很低,上升不明显,病毒滴度处于较低水平,上升趋势也不明显;12~24 h期间,病毒RNA和病毒滴度开始明显上升;24~72 h期间,病毒RNA急剧上升,而病毒滴度在24~48 h期间明显上升,48 h后趋于平稳,72 h时病毒滴度达到最高,为106.75TCID50/mL。GCRV 873株感染CIK后12 h时出现少量细胞聚集;24 h时,细胞聚集增多,出现空斑现象;36 h时,空斑变大;48 h时,细胞大量脱落;72 h时,细胞基本已脱落,只有零星细胞附着于板上(图5)。
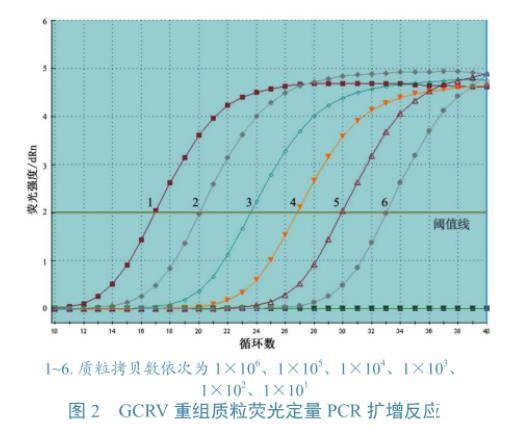
图2 GCRV重组质粒荧光定量PCR扩增反应
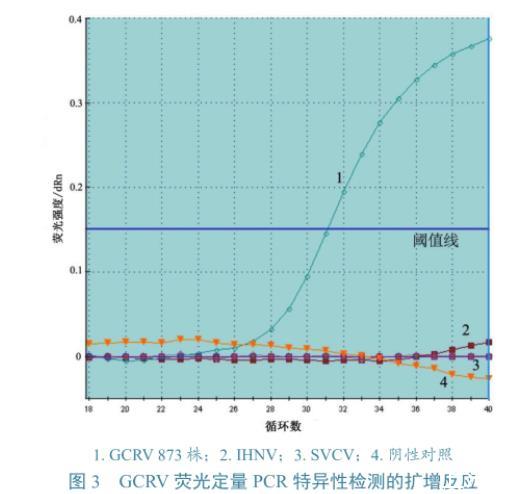
图3 GCRV荧光定量PCR特异性检测的扩增反应
2.3 GCRV 873株在草鱼体内的复制情况
GCRV 873株感染草鱼后,其肝、脾、肾中的病毒RNA拷贝数变化如图6所示。3个组织中的病毒RNA均呈先升后降趋势,其中肝脏中的病毒RNA峰值位于感染后3 d,脾脏和肾脏峰值位于感染后2 d。3种组织中,病毒RNA峰值最高的为肾脏,肝脏次之,脾脏最低;肝脏、肾脏和脾脏中的病毒RNA分别在至感染后7、6、4 d时下降到趋近于零的水平。
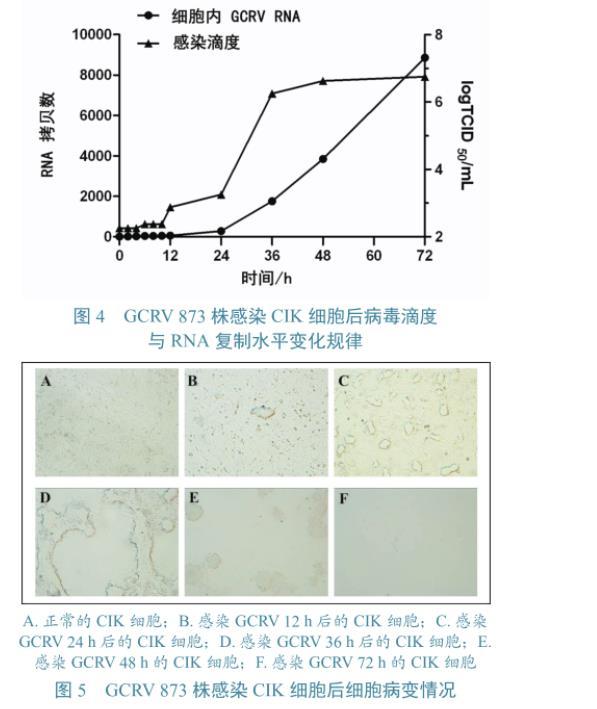
图4 GCRV 873株感染CIK细胞后病毒滴度与RNA复制水平变化规律 图5 GCRV 873株感染CIK细胞后细胞病变情况
3讨论
本研究从病毒RNA拷贝数、子代病毒含量及细胞病变3个方面综合分析GCRV在CIK细胞中的增殖过程。试验结果表明,GCRV在感染CIK细胞的早期病毒RNA拷贝数及病毒滴度均处于较低水平,说明这一时期的病毒多为残余的原代病毒及最先释放出来的子代病毒;24~72 h期间,病毒RNA水平迅速上调,呈现对数增长,而病毒滴度在24~48 h期间呈明显上升,之后趋于平稳;对照48 h的细胞CPE发展完全,说明细胞被完全裂解,胞内病毒得以释放,使病毒滴度趋于平稳。曾令兵等利用组织培养微量滴定系统,研究GCRV-854毒株在CIK细胞上的繁殖过程,其动态曲线与本研究相似,但出现快速增殖的时间较本研究晚,说明不同毒株感染细胞的能力存在差异。邹桂平等在电镜下观察到GCRV在感染CIK细胞4 h以内出现脱去部分外层衣壳的不完整病毒颗粒,感染8 h时浆胞内出现大量亚病毒颗粒,无外层蛋白结构,感染12~16 h后出现成熟的病毒粒子。丁清泉等发现GCRV在感染细胞12 h后即开始增殖,24~72 h大量增殖,使细胞产生典型的细胞病变效应,5 d左右达到最大增殖,此时病毒的滴度最高,以后逐渐平缓。分析病毒在细胞中的增殖过程,不仅反映出病毒的生长特性,还指示出收获细胞培养的病毒材料的最佳时间。
应用Real-time PCR技术,实时监测GCRV在草鱼体内的病毒拷贝数,为精确分析病毒在鱼体中的增殖过程提供基础数据。本试验采用的毒株GCRV 873株为1型。该毒株在体外细胞中培养可使细胞产生明显病变,但是草鱼感染后未出现发病死亡情况,而其肝、脾、肾中的病毒RNA水平出现短期上升后下降到对照水平的结果,也正与无发病死亡情况相符。殷亮研究发现GCRV HZ08株在感染稀有鮈鲫后,脾脏和肾脏中的病毒RNA水平呈现先升后降趋势,与本研究结果一致。丁清泉等发现经人工感染GCRV的鱼体肾脏组织细胞内,存在无外衣壳的未成熟病毒。毛树坚等观察草鱼出血病的病理切片发现在肝脏、肌肉、肾脏、脾脏、鳃等鱼体组织中含有大量的病毒颗粒。在本研究对比3种组织中病毒RNA的含量,发现肾脏中的含量最高,提示在取样检测时,可优先考虑肾脏组织。
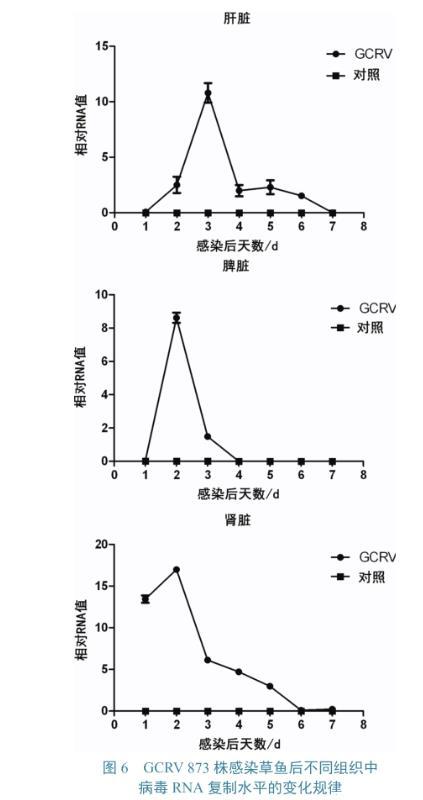
图6 GCRV 873株感染草鱼后不同组织中病毒RNA复制水平的变化规律
4结论
本研究显示:GCRV在感染CIK细胞12 h后病毒RNA和病毒滴度开始上升;24~72 h期间,病毒RNA急剧上升,病毒滴度则在24~48 h内明显上升,48 h后趋于平稳,至72 h,病毒滴度达到最高,为106.75TCID50/mL。在感染CIK后36 h,所有细胞均已感染,72 h时,细胞基本上脱落。GCRV感染草鱼后,均能在其肝、脾、肾中检测到病毒,且病毒RNA均呈先升后降趋势;肾脏中的病毒RNA含量最高,因此应优先采集肾脏组织进行检测。
相关新闻推荐
2、水貂李氏杆菌的分离鉴定、生化试验、药敏实验、毒力试验——结果、讨论