酿酒酵母和耐热酵母属间原生质体融合频率及方法
酿酒酵母(Saccharomyces cerevisiae)396是利用甘蔗糖蜜生产酒精的生产菌株,假丝酵母(Candidasp.)C6是我们从云南温泉底泥中筛选到的一株能在45℃生长良好的耐热酵母,我们应用原生质体融合技术进行了两菌株属间融合的研究。通过亚硝基胍(NTG)诱变得到的营养缺陷型菌株396(arg-)和C6(lys-),其融合频率为0.91x10-5。从检出的融合子中挑选6株进行考核、其细胞体积平均为亲株的1.3倍,DNA含量平均为亲株的1.6倍,培养特征、形态、大小和生理生化特征表现不一,特别是糖类的同化试验。除F2、F14外,其他4株可以排除异核体形成的可能性。比较了在28℃,40℃和45℃培养条件下出发亲株396、C6、直接亲株396(arg-)C6(1ys-)和融合子F1、F7、F12、F13的生长曲线、基质利用率和乙醇产率等,得到一株在40℃培养条件下糖的利用率为94.3%、乙醇产量为59.7g/L的属间融合株F13。
自1977年Ferenczy等将原生质体融合技术用于酵母以来,关于酵母菌的种内融合和种间融合有不少报道。1978年Provos+等又报道获得了热带假丝酵母(Candida tropicalis)和扣囊复膜孢酵母(Saceharo-mycopsis fibuligera)的属间融合子。1984年品川早苗将酿酒酵母(Saccharomyces cerevisiae)N1,一株高产乙醇但不耐热的菌株与耐高温的菌株TJ1融合得到AM 12,可以在40℃高产酒精。
本文对生产酒精的酿酒酵母(S.cere-visiae)396和能在45℃生长良好的耐热酵母(Candida sp.)C。的属间原生质体融合进行了研究。
材料与方法
(一)菌株
原始出发菌菌株:酿酒酵母(Saccha-10nyces cerevisiae)396是云南省建水糖厂提供的甘蔗糖蜜酒精生产菌株;假丝酵母(Candida sp.)C。是我们从云南温泉的底泥中分离到的一株能在45℃生长良好,并能发酵糖产生乙醇的耐热酵母。融合用直接亲株则是以上两菌株经NTG诱变后得到的营养缺陷型菌株396(arg-)和C。(lys-)。
(二)培养基
完全培养基(YPD)(%):酵母粉(上海酵母厂)0.5,蛋白胨1,葡萄糖2,固体加琼脂2;高渗完全培养基(YPD)即YPD加17%蔗糖;基本培养基(YNB)采用Wi-加17%蔗糖;基本培养基(YNB)采用Wi-cherham培养基;融合子检出培养基(YNBS)即YNB加蔗糖17%。
酒精发酵用培养基:甘蔗糖蜜(云南省建水糖厂提供)17Bx°,硫酸铵0.2%,磷酸0.1%,pH4.5。
(三)试剂
0.2mol/L磷酸缓冲液(PB),pH5.8;亚硝基胍(NTG)-PB诱变剂中NTG浓度
1mg/ml,使用剂量250ug/ml;制霉菌素(NY)诱变浓缩剂使用剂量150μg/ml,用二甲基甲酸胺溶解NY;用于耐热酵母C_6left(1,s^{-}right)的预处理剂是Tris 100mmol/L,DTT(二硫基苏糖醇)5 mmol/L,EDTA 5 mmol/L;用于酿酒酵母396(arg-)的预处理剂是Tris 100mmol,巯基乙醇0.5%;脱壁酶液是中科院生物物理所提供的蜗牛酶0.5%(1%)加0.7mmol/L甘露醇溶于磷酸缓冲液(PB)中,使用前配制并用G 6细菌漏斗过滤除菌;助融剂为35%聚乙二醇(PEG MW.6000)-10mmol/L氯化钙溶液。
(四)方法
采用文献和的方法通过NTG诱变出发菌株后,又采用文献的方法用NY浓缩出营养缺陷型菌株,最后用Fin-cham等的方法进行菌株营养缺陷型的鉴定;原生质体的制备、再生和融合的方法参考文献和;DNA的测定参考文献的方法,用2g重结晶二苯胺加入100ml冰乙酸和10ml高氯酸,使用前加入1.6%的乙醛溶液1ml,30℃保温14h。采用酶解法破壁后提取DNA,但C。有少数细胞壁未破,异核体的检测参考文献的方法,乙醇含量的测定采用气相色谱内标标准曲线法,内标物用甲乙酮;糖量的测定用3,5-二硝基水杨酸定糖法。
(五)参数的计算
1.突变率(a):a=left(M-M_0right)/left(N_0-right.N),式中M。:起始突变数,一般为0,M:最后的突变数,No:起始活细胞数,N:最后活细胞数。
2.原生质体形成率和再生率:原生质体形成率(%)=(A-B)/Ax100%;原生质体再生率(%)=(C-B)/(A-B)x 100%;A:蜗牛酶处理前菌落数(YPD平血);B:蜗牛酶处理后未脱壁细胞形成的
菌落数(YPD平血);C:蜗牛酶处理后未脱壁细胞形成的菌落数加原生质体再生细胞菌落数(YPD平血)。
3.原生质体融合频率(Ff):

结果与讨论
(一)营养缺陷型标记菌株的获得
用250μg/mlNTG诱变1h,150μg/ml NY浓缩后检出精氨酸缺陷型菌株396(arg-)、赖氨酸缺陷型菌株和亮氨酸缺陷型菌株Cσ(lcu-)(表1)。
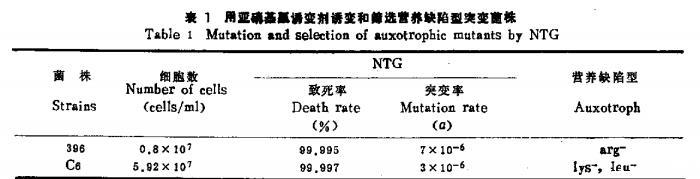
396(arg-)和Cσ(lS-)没有检出回复突变株,而C_8left(right.leuleft.^{-}right)的回复突变率为10^{-4}。从细胞形态上观察396(arg-)和C_8left(1~s^{-}right)都与原出发菌株相似,而C_6(leu-)不论在YPD固体培养基上或液体培养基中均呈簇生的假菌丝型,不宜用于融合。
(二)原生质体的制备和融合
营养缺陷型菌株静止培养14h,离心收集菌体,30^{circ}C预处理20min后进行脱壁。由于0.5%和1.0%的蜗牛酶对396(arg-)的脱壁率分别为78.04%和77.77%(血球计数板计数),差异不太大,所以我们采用0.5%蜗牛酶。396(arg-)的细胞浓度为4.05times 10^6 cells/ml,原生质体形成率为92.86%,再生率为4.83%。而Ce(lys-)的细胞浓度为2.0x10°cells/ml,原生质体形成率为84.57%,再生率为7.75%。C。(lys)的细胞破壁较396(arg-)困难一些。
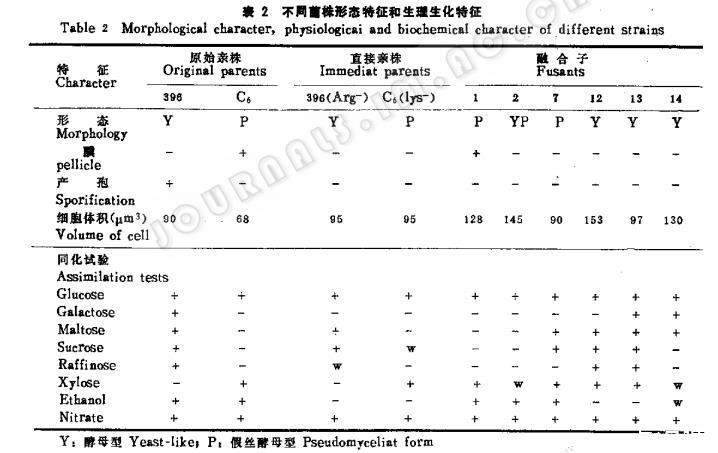
营养需求看除了F,在YNB上有时生长,有时不生长,而在补充了精氨酸的YNB平皿上完全不生长外,其他均为原养型菌株,融合子与原始亲株和直接亲株的生物学特性比较见表2。DNA含量的比较见表3。
(三)融合子的生物学特性
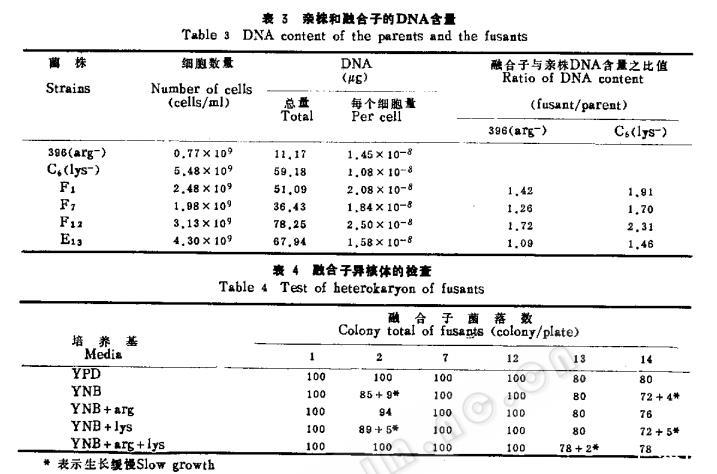
随机挑出6株融合子进行考核。从其从表2和表3可以看出融合子的培养特征形态,大小、每个细胞的DNA含量和生理生化特征表现不一,特别是糖类同化更是多样,其细胞体积平均为亲株的1.3倍,DNA含量平均为亲株的1.6倍。
对融合子异核体的检查结果(表4)除F2和F14外,其他4株F1、F7、F12和F13可以排除异核形成的可能性。
(四)融合子的耐热性和乙醇产量
1.在不同温度条件下亲株和融合子的生长曲线:出发亲株、直接亲株和融合子在YPD液体培养中静止培养在28℃、42℃和45℃,72h的生长曲线(图1)表明营养缺陷型菌株的耐热性比出发菌株显著下降,而回到原养型的融合子耐热性虽然各株不一,但也都比原出发亲株略差。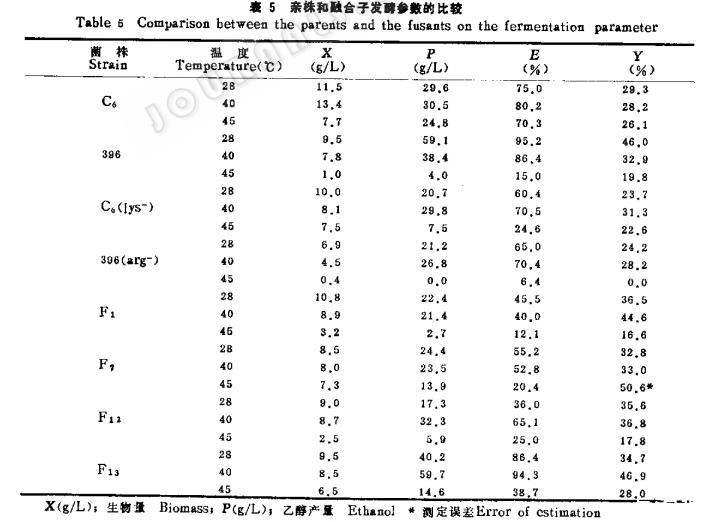
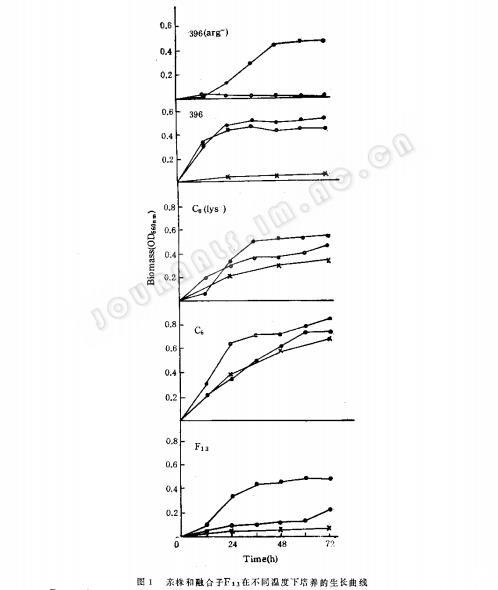
2.不同温度条件下基质消耗和乙醇产量的比较:各类型菌株在28℃、40℃和45℃培养条件下,利用甘蔗糖蜜发酵乙醇的各种参数(表5)表明,营养缺陷型菌株396(arg-)在不同温度下,乙醇的产率和基质的利用都比原出发亲株:396低,经过与耐热菌株C。(1s-)融合后得到的融合子,只有F13一株乙醇产率和基质利用率比较理想。在40℃和45℃下培养,乙醇产量均高于原生产菌株396。
相关新闻推荐
1、从细菌/真菌对数生长曲线到降解率:微生物适应性与芳烃去除的相关性