空肠弯曲菌噬菌体生物学特性测定及生长曲线绘制(三)
2.5噬菌体vB_Cj_QDYZ功能注释
噬菌体vB_Cj_QDYZ全基因组的功能注释如图3所示,全基因组一共有165个ORF,3个tRNA。其中有65个ORF编码已知功能,另外100个编码假设蛋白。对65个具有已知功能的ORF分类,主要有4种功能的蛋白:噬菌体结构蛋白、裂解蛋白、包装蛋白、与噬菌体复制和代谢有关的蛋白。具体的注释结果见附表1(https://doi.org/10.13995/j.cnki.11-1802/ts.035791)。
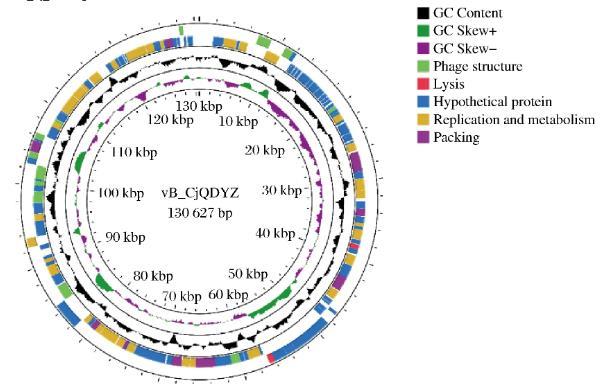
图3噬菌体vB_Cj_QDYZ基因组功能注释
噬菌体结构蛋白有:ORF10(主要衣壳蛋白),ORF131(衣壳组装蛋白),ORF11和ORF34(头部完成蛋白),ORF133(颈部蛋白),ORF13和ORF167(尾鞘稳定剂和完成蛋白),ORF90、ORF91和ORF136(尾管蛋白),ORF113(尾鞘蛋白),ORF100(尾丝蛋白)。其中,尾管蛋白是噬菌体编码尾部的蛋白,与吸附宿主菌密切相关。尾丝蛋白是噬菌体特异性识别宿主的物质基础,噬菌体的尾丝蛋白在特定微生物检测净化等方面具有重要意义。
噬菌体裂解相关蛋白有2个:ORF66(内溶素)和ORF85(尾部溶菌酶)。噬菌体可能是通过表达这两种蛋白来裂解宿主菌的。
噬菌体包装蛋白:ORF107(末端酶大亚基),ORF134(末端酶小亚基)。噬菌体包装蛋白主要在噬菌体包装时起重要作用。其他蛋白包括受体结合蛋白:ORF1、ORF2、ORF3、ORF4,与噬菌体结合细菌表面受体相关。
与噬菌体复制和代谢有关的蛋白有:ORF6、7(重组核酸内切酶),ORF8(转录调控因子),ORF37(RNA连接酶),ORF48(聚合酶),ORF55(重组和修复蛋白),ORF63(脱氢酶),ORF68(拓扑异构酶),ORF97(氨基转移酶家族蛋白),ORF101(核糖核酸酶),ORF158(DNA聚合酶),ORF162(胸苷激酶),ORF164(DNA引物酶亚基),ORF12、ORF54和ORF87(归巢核酸内切酶),ORF71、ORF144和ORF145(解旋酶),ORF61(dUTP焦磷酸酶),ORF153(核酸外切酶)。其中,重组和修复蛋白的功能是将噬菌体基因组整合到宿主菌上。归巢核酸内切酶参与DNA内含子的水平转移和基因表达的调控。解旋酶是一类解开氢键的酶,一般在噬菌体DNA复制过程中起到催化双链DNA解旋的作用。焦磷酸酶,参与DNA合成。核酸外切酶,具有从分子链的末端顺次水解磷酸二酯键而生成单核苷酸作用的酶。
2.6噬菌体vB_Cj_QDYZ比较基因组分析
噬菌体vB_Cj_QDYZ与空肠弯曲菌噬菌体CP39、CJLB-10、CP30A、CPX、CP8的同源性分别为98.30%、94.77%、94.61%、97.69%、97.82%,比对覆盖率分别为95%、91%、92%、92%、92%。弯曲菌噬菌体分为2个属Fletchervirus和Firehammervirus。与vB_Cj_QDYZ相似度最高的噬菌体来自Eucampyvirinae亚科,Fletchervirus属。初步推测空肠弯曲菌噬菌体vB_Cj_QDYZ可能是来自Eucampyvirinae亚科,Fletchervirus属的新成员。
2.7噬菌体vB_Cj_QDYZ系统进化分析
如图4-A所示,基于全基因组序列构建的进化树结果表明,噬菌体vB_Cj_QDYZ与CampylobacterphageCP39具有最近的亲缘关系。NCBI检索发现,CampylobacterphageCP39是Eucampyvirinae亚科、Fletchervirus属的成员。研究表明CP39是肌尾噬菌体。因此推测vB_Cj_QDYZ是肌尾噬菌体(Myoviridae)科、Eucampyvirinae亚科、Fletchervirus属的成员。
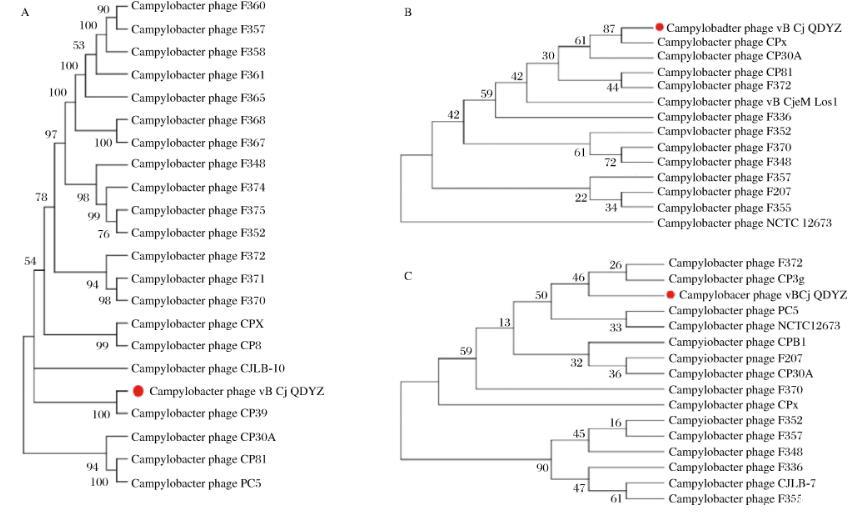
A-基于全基因组构建的进化树;B-基于内溶素序列构建的进化树;C-基于主要衣壳蛋白构建的进化树
图4噬菌体vB_Cj_QDYZ进化关系分析
如图4-B所示,噬菌体vB_Cj_QDYZ的内溶素和CampylobacterphageCPX的内溶素相似度最高;如图4-C所示,vB_Cj_QDYZ的主要衣壳蛋白与CampylobacterphageCP39的最相近。标尺代表遗传距离,遗传距离越短代表亲缘关系越近。明显看出,图4-B和图4-C的进化树标尺较长,遗传距离较大。因此推断噬菌体vB_Cj_QDYZ可能为一株新的空肠弯曲菌噬菌体。
2.8噬菌体在牛奶中的抑菌效果
如图5所示,加入空肠弯曲菌噬菌体vB_Cj_QDYZ后,在正常环境下放置4 h后,通过平板计数法测定,哥伦比亚血平板上的空肠弯曲菌菌落总数从初始的1.0×107CFU/mL降低至1.17×105CFU/mL,降低了2个lg CFU/mL,充分说明噬菌体vB_Cj_QDYZ在牛奶中的杀菌率为99%。牛奶是容易被弯曲菌污染的食品,选择牛奶作为研究对象具有实际参考价值。综上所述,噬菌体vB_Cj_QDYZ具有良好的抗菌潜力。

图5噬菌体vB_Cj_QDYZ在牛奶中的抑菌效果
3结论与讨论
本研究以空肠弯曲菌2206为宿主菌,从鸡粪中分离出一株空肠弯曲菌噬菌体,将其命名为vB_Cj_QDYZ。噬菌体在双层平板上可形成清晰透亮的噬菌斑,这说明噬菌体具有很强的裂解能力。透射电镜形态表明,噬菌体有一个正二十面体的头部(直径约100.81 nm)和一条可伸缩的尾部(长度约65.90 nm),具有弯曲菌噬菌体典型的终端泡和空头结构,属于有尾噬菌体目,肌尾噬菌体科。大多数弯曲菌噬菌体属于肌尾噬菌体。
生物学特性结果显示,噬菌体具有良好的热稳定性和pH稳定性,表现出噬菌体在某些恶劣环境下具有一定的适用性。噬菌体vB_Cj_QDYZ在30~60℃,pH值为3~12的环境下效价较稳定,这为噬菌体在实际家禽生产中的防控提供了理论依据。空肠弯曲菌主要寄居在鸡等家禽的肠道中,鸡的正常体温是39.6~43.6℃,鸡肠道的pH值主要为2~4,这说明噬菌体vB_Cj_QDYZ可以在鸡的肠道中存活且保持较高效价,具有广泛的应用范围。噬菌体vB_Cj_QDYZ的最佳感染复数为0.01,充分表明了噬菌体的侵染能力较强。一步生长曲线表明,噬菌体的潜伏期是60 min,裂解期是150 min,爆发量是45 PFU/cell。因此,空肠弯曲菌噬菌体vB_Cj_QDYZ具有较短的潜伏期和较强的裂解能力。此外,加入不同感染复数的噬菌体显著抑制了空肠弯曲菌的生长,表现出噬菌体vB_Cj_QDYZ具有很强的裂解能力。噬菌体vB_Cj_QDYZ在牛奶中的杀菌率为99%,实际应用价值较高,具有开发为新型空肠弯曲菌抗菌剂的潜力。
基因组学研究有利于从基因水平更深入了解空肠弯曲菌噬菌体。噬菌体vB_Cj_QDYZ基因组为双链DNA,基因组全长为130 627 bp,G+C含量为26%。基因组大小位于130~140 kbp,根据ICTV对弯曲菌噬菌体的最新分类,噬菌体vB_Cj_QDYZ被命名为CP8 virus,且属于Ⅲ组噬菌体,特异性受体是荚膜多糖,对空肠弯曲菌具有较强的裂解性能。Ⅲ组噬菌体是目前抑菌效果最好的空肠弯曲菌噬菌体。噬菌体一共有165个ORF,3个tRNA。其中65个ORF具有功能。此外,以全基因组序列构建的进化树显示,噬菌体vB_Cj_QDYZ与CampylobacterphageCP39具有最近的亲缘关系以主要衣壳蛋白和内溶素序列构建的进化树表明,噬菌体与其他的空肠弯曲菌噬菌体遗传距离较远,推断噬菌体vB_Cj_QDYZ属于Fletchervirus属的新成员。此外,噬菌体vB_Cj_QDYZ不含有抗生素耐药基因、毒力基因、过敏原基因等,安全性较高,适合于实际应用生产。
本研究分离纯化一株裂解能力较强、稳定性良好、安全性较高的新型空肠弯曲菌噬菌体,能够有效抑制空肠弯曲菌的生长,且特异性较强,具有开发为新型空肠弯曲菌抗菌剂的潜力,丰富了空肠弯曲菌噬菌体的资源,为防治空肠弯曲菌污染提供理论基础和技术指导。
相关新闻推荐
2、VgrS受体细菌组氨酸激酶在NYG培养基中的的生长曲线图