量化人类肠道菌种对艰难梭菌长期生长的影响(五)
根据推断的gLV参数,两种交互网络都显示出频繁的负交互作用,符合之前的研究发现(图4B和4C)。与祖先株(18%)相比,进化株(25%)存在更多的正交互作用。值得注意的是,15%的推断交互作用具有不一致的符号。祖先株和进化株艰难梭菌株之间的种间交互系数显示出中度的相关性(皮尔逊R=0.59,p=1.8E?06;图4D和S12D)。相比之下,作为参考,不同遗传背景的艰难梭菌株之间种间交互作用的差异不频繁(所有艰难梭菌株对之间的皮尔逊R值为0.81–0.89,p=3.4E?14至7.6E?20)。这一发现突显了艰难梭菌代谢基因中两个关键单点突变对人体肠道微生物种间交互作用的重大影响。例如,CS和CH对祖先株的抑制作用显著大于进化株。由于CS和CH在我们的培养基中显著消耗了脯氨酸(图S6B),因此与艰难梭菌代谢生态位的改变相比,祖先株可能与CS和CH之间的资源竞争程度高于进化株。
艰难梭菌的演化适应间接影响了其他群体成员之间的配对交互作用(图4B–4D),这表明网络中可能存在更高阶交互作用(HOI)。为了直接捕捉潜在的HOI,我们使用了一种修改的gLV模型,其中包含群体成员之间的配对和第三阶交互作用(gLV-HOI)进行数据拟合(图S12E、S13C和S13D;STAR方法)。通过20倍交叉验证,gLV-HOI显示出高预测性能(皮尔逊R=0.93–0.98,p<10E?05),但未能在基于相关系数z检验的标准gLV模型中获得更优表现(表S12;图S12C)。这表明群落动态主要由二阶交互作用驱动。gLV-HOI的参数数量大于标准gLV模型,因此使用有限实验数据进行参数估计更加挑战。为了进一步研究HOI项的贡献,我们使用套索回归(STAR方法)评估了gLV-HOI的预测性能。套索回归使用L1正则化,这会惩罚参数估计的绝对值总和(L1范数),促进模型的稀疏性。相比之下,贝叶斯推断中的正态先验引入L2正则化,惩罚参数与先验均值之间的平方差,从而实现参数估计的平滑收缩,而不是驱动参数趋于零。在不显著改变预测性能的情况下,正则化系数的最大值迫使所有HOI参数为零(图S14A–S14D)。这意味着在该数据集上,gLV模型的预测性能不会因HOI项而改善。然而,我们不能排除gLV-HOI在捕捉社区潜在HOI交互作用方面不够灵活的可能性。总之,艰难梭菌关键的演化适应导致其代谢生态位的显著改变,给人体肠道群落配对交互作用的推断带来了重大改动(图4B–4D)。
演化适应的艰难梭菌菌株在哺乳动物肠道中减轻了疾病严重程度
氨基酸如脯氨酸已被证明能够调节艰难梭菌毒素的体外生成,并影响哺乳动物肠道的定植。为了确定艰难梭菌的关键演化适应是否会改变毒素的生成,我们通过酶联免疫吸附检测法(enzyme-linked immunosorbent assay,ELISA)在培养24小时后进行了毒素表达分析。演化适应的艰难梭菌菌株显示出显著较低的毒素浓度和产量(浓度与OD600之比),相比于祖先菌株,在多种氨基酸浓度下均如此(Figures S15A和S15B)。
为了研究演化适应的艰难梭菌菌株是否影响疾病的严重程度,我们利用小鼠模型,将演化适应的或祖先的艰难梭菌菌株通过口服灌胃引入无菌小鼠体内,实验中部分小鼠没有人体肠道菌群(见图5A)。具体而言,小鼠分别通过口服灌胃接种单一菌群(PV),另一部分小鼠则接种由七种人体肠道菌群组成的菌群(CS,CH,DP,BT,PV,BU,CA),这些菌群在我们的长期培养实验和菌群特性分析中均有应用(见图1和4)。这些菌群在挑战前的8天被引入,旨在促进小鼠免疫系统的发育和菌群定植。在单独挑战艰难梭菌的小鼠组和带有七种菌群的小鼠组中,相较于祖先菌株,演化适应的艰难梭菌菌株表现出显著较低的丰度和毒素浓度。这种艰难梭菌的定殖减少可能导致毒素生成较少,因为毒素产量(毒素量除以艰难梭菌菌落形成单位[colony-forming unit,CFU])在两个组之间相似(见图S15D)。这种与体外实验中的毒素产量不一致(见图S15B)的现象表明,毒素在体内和体外的调控机制存在差异。为了更全面地探讨祖先或演化适应的艰难梭菌菌株的影响,我们聚合了所有处理组的数据,分析微生物群落组成的变化(见图S15C)。在所有微生物群落条件下,挑战演化适应的艰难梭菌的小鼠显示出更高的存活率,并显著低于挑战祖先菌株的小鼠,相对体重减轻尽管所有小鼠在最初几天体重均显示出下降趋势(见图5B–5D和S15C)。
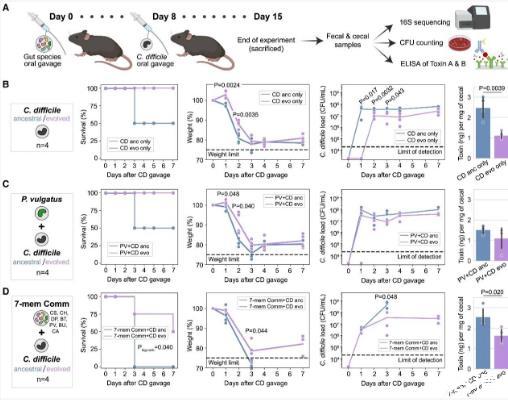
图5|演化适应的艰难梭菌菌株缓解了小鼠的疾病严重性
(A)小鼠实验示意图。小鼠在引入原始或演化适应的艰难梭菌菌株前,分别被挑战于没有任何肠道菌群干预(仅艰难梭菌组)或通过PV菌株或7种肠道菌群组合(CS、CH、DP、BT、PV、BU和CA)进行口服灌胃8天。
(B–D)小鼠在仅接受艰难梭菌(n=4)(B)、PV与艰难梭菌(n=4)(C)以及7种肠道菌群与艰难梭菌(n=4)(D)口服灌胃后,存活百分比、初始体重百分比、艰难梭菌丰度以及艰难梭菌毒素浓度(每mg盲肠内容物)。在存活率上,log-rank检验显示了显著的p值。在初始体重百分比上,线条表示该组所有小鼠的平均值。体重下降至75%以下的小鼠会被淘汰。在艰难梭菌丰度方面,通过CFU计数法确定值,该方法使用了盲肠内容(存活)和盲肠内容(死亡)进行艰难梭菌选择性培养基的检测。在盲肠内容中的艰难梭菌毒素浓度方面,数据以平均值±标准差表示。由于在多个治疗组中,小鼠在第3天死亡,因此在第4天之后的数据点比总的实验小鼠数量少。因此,体重(%)和艰难梭菌丰度的统计检验仅在数据完整的第1到第3天进行。对于体重(%)、艰难梭菌丰度和艰难梭菌毒素浓度的图示,使用两侧的未配对t检验显示了显著的p值。部分图表由Biorender生成。
PV单菌群显著降低了祖先艰难梭菌菌株的毒素浓度,使其与演化菌株达到相似水平(见图5B、5C和S15E)。同时,PV显著增加了演化菌株艰难梭菌菌株在某些时间点的定殖(见图S15F),导致祖先菌株和演化菌株的艰难梭菌菌株在定殖方面达到相似水平(见图5B和5C)。根据16S rRNA基因测序,PV在挑战演化菌株的小鼠中相对丰度较高,而与祖先菌株相比(~77%对60%,7天后)(见图S15G)。由于祖先菌株和演化菌株的绝对丰度在CFU计数法下相似(见图5C),这一结果表明,PV在携带演化菌株的小鼠中的丰度高于携带祖先菌株的小鼠。因此,这些数据表明,PV和艰难梭菌菌株之间存在双向互动,影响小鼠肠道中的定殖能力和毒素生产。
在口服七种肠道菌群的小鼠中,存活率显著低于仅携带艰难梭菌和PV单菌群的小鼠组(见图5D)。这表明,七种肠道菌群显著增加了艰难梭菌的疾病严重性。挑战后第三天,在携带七种肠道菌群的小鼠中的祖先艰难梭菌菌株丰度显著高于其他组(p值=0.02)(见图5B–5D)。PV在携带祖先或演化菌株的小鼠中显示出最高的相对丰度(~50%,接种几天后,见图S15H)。由于PV单菌群定殖未显著改变疾病严重性,因此携带七种肠道菌群的小鼠相比其他组观察到的疾病严重性增加可能是由其他菌群的存在、PV与艰难梭菌的交互作用变化或宿主-微生物交互作用导致的。总之,在所有条件下,与祖先菌株相比,演化菌株艰难梭菌菌株在小鼠肠道中显示出显著降低的定殖能力和疾病的严重性(见图5B–5D和S15C)。
相关新闻推荐
3、焦化污染场地中萘降解菌株AO-4鉴定、生长、降解机理及环境条件带来的影响(二)