从儿童到成人:面部皮肤菌群成熟过程中的菌株建立与稳定规律(三)
谱系共享由多细胞传播介导
为深入了解成功传播事件中涉及的传播方向及细胞数量,我们分析了基于分离株构建的谱系系统发育树(图3B–4F)。当个体分离株的最近共同祖先(MRCA)明显晚于另一位个体时,可据此推断传播方向。然而,在具备足够分离株的谱系中,仅有少数(40个谱系中18个)满足这一条件(图3E;详见STAR方法)。
在这些可推断方向的传播事件中,大多数是由父母传播给子女(痤疮丙酸杆菌中为9/10,表皮葡萄球菌中为9/13;见图3B、4C)。若系统发育树呈现复杂拓扑结构,往往难以确定方向性,这可能是由于涉及多个个体、多个基因型的传播,或两者共同作用(图3E)。在家庭成员之间共享的谱系中,我们观察到痤疮丙酸杆菌和表皮葡萄球菌均以多细胞传播为主。估算显示,每个谱系在每位接收个体中通常涉及至少两个不同基因型(即多个细胞)的传播,最多可达11个。
由于同一基因型的多个细胞传播无法区分,这一估算仍偏保守。此外,在痤疮丙酸杆菌中,随着分离菌落数量的增加,推断出的传播基因型数量也相应增多。虽然我们无法判断这些细胞是同时还是分时传播,但整体结果表明,在人与人之间的传播中,单细胞瓶颈现象较为罕见。对于痤疮丙酸杆菌,多细胞传播的普遍性与我们此前的研究相符,即:每个皮脂毛囊单独定植,形成类似“群岛”的空间结构,有助于多个基因型在不直接竞争的条件下长期共存。相比之下,表皮葡萄球菌在皮肤上的微观空间分布仍不清楚,目前尚不清楚其是否存在支持多基因型共存的解剖学基础。
此外,痤疮丙酸杆菌和表皮葡萄球菌的系统发育结构也存在显著差异:前者具有更“梳状”的树形结构,突变不共享的比例更高,而后者谱系之间的差异相对较小,这可能反映出表皮葡萄球菌菌群在皮肤上混合更充分。综上所述,这两种菌在传播过程中均未出现明显的单细胞瓶颈现象,提示其在宿主体内可能存在空间结构化的生态环境,从而有利于多个传播基因型的稳定共存。
表皮葡萄球菌(S.epidermidis)谱系持续被获取,并随时间逐渐稳定
接下来,我们通过比较连续采样,进一步探究个体体内谱系的动态变化。对于表皮葡萄球菌(S.epidermidis),我们在较短的时间间隔内(中位时间为0.43年,范围0.39–1.0年)观察到大量谱系更替事件。与相对稳定的痤疮丙酸杆菌相比,表皮葡萄球菌在不同时间点之间丢失的谱系比例显著更高,其谱系层级群落的总丰度下降也更为明显(图4A和5B)。这种动态性也在群落水平上表现为快速的变化,尽管在任意一个时间段内,个体的谱系并未发生完全更替(图4C)。尽管此前有研究(如Oh等人)认为表皮葡萄球菌在该时间尺度上较为稳定,但我们对该研究数据的重新分析显示,与本研究观察到的结果一致,表皮葡萄球菌实际上呈现持续更替的状态,其谱系丢失速率相似。
此外,这种波动不仅出现在儿童中,也同样发生在父母个体中,说明表皮葡萄球菌谱系在整个生命过程中都处于不断获取与流失的动态状态。
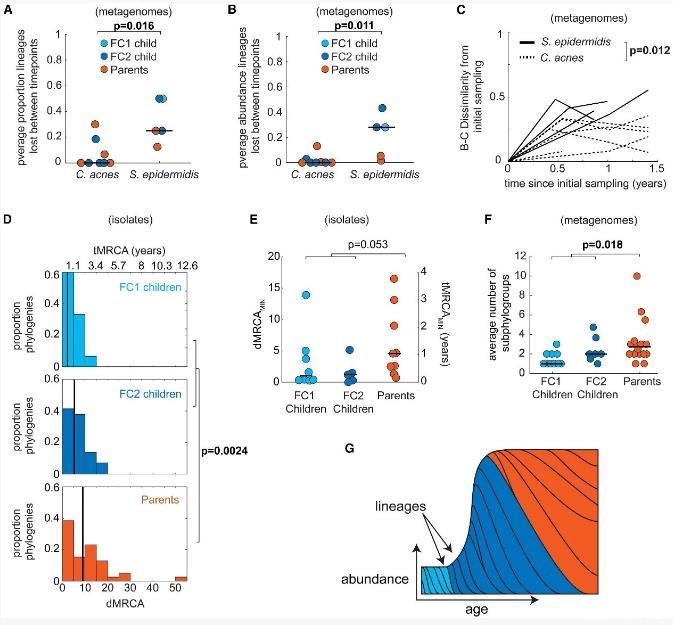
图4|表皮葡萄球菌(S.epidermidis)的谱系在整个生命过程中不断获得与丢失我们在多时间点具备足够宏基因组覆盖度的受试者中,分析了谱系水平的稳定性。(A)在谱系水平上,表皮葡萄球菌的稳定性远低于痤疮丙酸杆菌。(B)由于低丰度谱系接近检测下限,可能导致谱系更替的假象,我们评估了不同时间点之间丢失谱系的总丰度。结果显示,表皮葡萄球菌丢失谱系的平均总丰度较大,而痤疮丙酸杆菌则几乎可以忽略不计。图中显示了每位受试者在所有时间间隔的平均值(痤疮丙酸杆菌n=8,表皮葡萄球菌n=5)。(C)在为期1.4年的观察期间,基于考虑丰度的Bray-Curtis不相似性指标,个体体内表皮葡萄球菌的谱系群落更替速度明显快于痤疮丙酸杆菌。图中显示了在至少两个时间点拥有超过70%谱系分配的受试者,且两次采样时间间隔至少6个月的个体(痤疮丙酸杆菌n=7,表皮葡萄球菌n=4;)。
(D)通过计算个体上各谱系的突变距离到最近共同祖先(dMRCA),并结合分子钟转换为最近共同祖先时间(tMRCA),进一步支持了表皮葡萄球菌的不稳定性。分析对象为每位受试者中至少含有5个分离株的谱系。表皮葡萄球菌谱系的中位年龄约为1.2年(n=83,最大12.2年)。谱系在父母身上的年龄显著大于儿童,尽管相较于成人寿命仍较年轻。这一年龄估计不是采样深度的假象,因为含有大量分离株的谱系往往具有较低的dMRCA。(E)各组受试者中新近定植的谱系(tMRCAMIN)年龄相近(FC1儿童n=8,FC2儿童n=5,父母n=9),提示新定植速率相似。(F)与较长的定植时间和相似的新定植率一致,父母体内的物种内丰富度高于儿童。物种内丰富度以表皮葡萄球菌的亚谱型数量衡量,涵盖无谱系数据的受试者(FC1儿童n=10,FC2儿童n=7,父母n=16)。
图中显示了每位受试者所有时间点样本的亚谱型平均数。仅分析亚谱型分配率超过70%的时间点样本(详见STAR方法)。(G)我们的观察结果支持这样一个模型:表皮葡萄球菌的谱系在个体间持续获得和流失,但在成人体内能够存续更长时间,成人具有更高的表皮葡萄球菌细胞承载能力[^30]。图中每组线条之间的区域表示谱系的丰度,不同颜色对应谱系被获得的时间窗口。所有p值均来自双侧秩和检验,使用Benjamini-Hochberg方法控制假发现率为5%,显著结果以粗体显示。组内水平或垂直线条表示中位数。为了用另一种独立方法验证我们关于表皮葡萄球菌(S.epidermidis)谱系不稳定性的推断,我们基于分离株基因组构建的谱系系统发育树,分析了每位个体上各谱系最近共同祖先(MRCA)的年龄。首先,我们测定了表皮葡萄球菌的突变积累速率,约为4.4次突变/基因组/年,与文献报道相符。然后,我们利用这一速率将谱系在个体上的遗传距离(dMRCA)转换为时间尺度(tMRCA)。正如纵向数据中观察到的动态不稳定性所预期,所有谱系的tMRCA都显著小于对应个体的年龄,最大值为12.2年,中位数为1.2年(图4D)。
虽然较低的tMRCA也可能源于近期的选择性扫荡或中性瓶颈导致的多样性丧失,但我们未发现与适应性进化相关的迹象,如平行进化、非同义/同义突变比率异常或时间点间的遗传扫荡。即使是在含有大量分离株的谱系中,也同样存在较低的tMRCA。这些结果表明,尽管表皮葡萄球菌在物种层面表现相对稳定,但其谱系在个体体内通常仅能存续数年。值得注意的是,父母体内的谱系tMRCA显著长于儿童(图4D;父母中位数约为2.1年,FC1和FC2儿童分别为0.53年和1.2年,p=0.0024),说明尽管谱系持续更替,个别谱系在成人体内更为稳定。这并非单纯因儿童出生时即被定植所致,因为儿童谱系的tMRCA仍明显低于其年龄。为了解不同年龄段新谱系定植速率的差异,我们考察了每个个体中最新获得谱系的年龄指标(tMRCAMIN),结果显示各组间无显著差异(图4E),支持表皮葡萄球菌在不同年龄个体中以类似速率获得新谱系,但谱系在成人中更稳定。基于上述两点——成人谱系稳定性更高和新谱系获得速率相似——我们推测成人体内谱系共存数量多于儿童。实际数据也显示,父母体内表皮葡萄球菌的物种内多样性显著高于儿童(图4F)。这与成人体内更高的菌群承载量(绝对丰度)降低了基因漂变,从而增强了稳定性相吻合。
综上,我们通过宏基因组的纵向数据和分离株系统发育分析两种独立手段,均得出一致结论:表皮葡萄球菌谱系从儿童期开始持续获得和流失,但在成人体内单个谱系的存续时间更长(图4G)。这一谱系层面的动态变化表明,终生的优先效应无法完全解释家庭成员间谱系共享不完全的现象(图3A)。
相关新闻推荐
1、肌苷对酸土脂环酸芽孢杆菌的生长抑制效果、对橙汁品质影响——讨论与结论
2、忍冬提取液对酸土脂环酸芽孢杆菌生长曲线、细胞形态和生物膜形成能力的影响(一)
3、基于ELISA和细菌生长曲线应用的结合定量检测肠炎沙门氏菌(二)