北京及非北京基因型结核分枝杆菌的生长曲线与基因表达差异
1995年,在中国北京及周边地区首次发现并报道MTB北京基因型菌株,将其命名北京型菌株。全球MTB临床分离株的13%是北京基因型菌株,并导致了全世界1/3的TB。与非北京基因型菌株相比,北京基因型菌株具有一定的选择优势,传播速度更快,毒力更强,往往更加致命。究竟什么原因赋予北京基因型菌株的这些特点还不完全清楚,被认为是相关特定分子的内在表达变化所决定。
研究结果显示,北京和非北京基因型菌株其生长曲线总体趋势都是标准条件下的OD值高于低氧及营养饥饿条件,正常标准培养条件下,由于氧气、营养充足、充分,适合MTB生长繁殖,OD值高;低氧条件下OD值低,MTB生长轻度受限,这种不利条件也是促进MTB持留生存的因素之一;营养饥饿条件下OD值最低,说明MTB生长速度最慢,繁殖速度降低以利于存活,也许这与MTB入侵人体被巨噬细胞吞噬后不被清除变为休眠菌机制相一致。与非北京基因型菌株相比,北京基因型菌株在低氧、营养饥饿的条件下OD值均高,即在不利条件下,北京基因型菌株生长繁殖速度快于非北京基因型菌株,说明其对低氧、营养饥饿不利条件耐受性较好,在压力条件下生存能力强,这也许是北京基因型菌株易流行的原因之一。是MTB什么内在的因素造成了这种不同,作者初步探索了MTB菌株的毒素-抗毒素mazEF系统与其生长繁殖的相关性。
毒素-抗毒素(TAS)由毒素蛋白和同源的抗毒素组成,mazEFTAS能够促进程序性细胞死亡,抑制细菌生长,诱导持留菌的形成,造成潜伏感染及提高细菌对环境压力的耐受性。TAS的作用机制取决于病原菌在不同生活时期的生物学特性,当受到不良环境或刺激后,打破了毒素抗毒素之间的平衡,抵抗力强的毒素被激活,作用于胞内靶分子,引起抑菌或杀菌效应。
Sat等对大肠杆菌进行mazEF基因敲除,通过比较其普通菌株和敲除株对不利环境如药物的反应,发现药物可以干扰或消耗不稳定的抗毒素,而使稳定的毒素MazF堆积,导致细菌的程序性死亡,因此推测mazEF可能是其程序性死亡进程中的关键因素。本研究通过敲除和过表达北京和非北京基因型菌株mazEF3、mazEF6和mazEF9基因后,观察各菌株在不同环境下的生长曲线,结果显示无论是北京还是非北京基因型菌株,其缺失突变株和过表达菌株的OD值均低于亲本株,提示mazEF3,6,9与MTB生长繁殖相关。缺失后,mazEF3,6,9不能发挥其应有的调节细菌生长繁殖作用,尤其是在不利的环境条件下,导致MTB菌数下降。过表达mazEF3、mazEF6和mazEF9基因,MTB菌数也降低,推测虽然抗毒素可以中和毒素,但是抗毒素不稳定,易降解,导致毒素增多、引起菌体死亡,或者是由于菌体内其他调节因素也参与了对其过表达调节作用的结果。究竟是何种原因导致的这种现象,需要进一步探讨。该结果也提示作者应进一步研究:在上述培养条件下,分别检测mazE3、mazE6、mazE9、mazF3、mazF6和mazF9对北京和非北京基因型菌株生长繁殖的影响。在mazEF系统mRNA表达水平上,北京基因型与非北京基因型菌株相比,mazEF3高表达;mazF3、mazF6、mazF9均高表达;mazE9低表达,差异均具有统计学意义(P<0.05)。
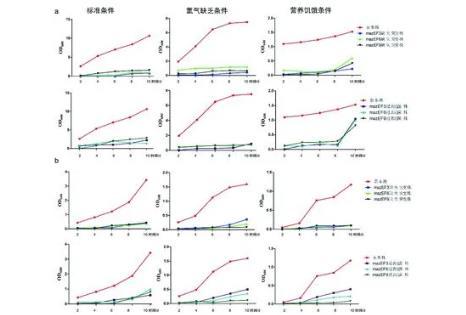
注:a. 北京基因型菌株及其缺失和过表达菌株在不同培养条件下的生长曲线;b. 非北京基因型菌株及其缺失和过表达菌株在不同培养条件下的生长曲线。图2 结核分枝杆菌及其mazEF3,6,9基因缺失和过表达菌株的生长曲线
有研究表明,分枝杆菌中Rv1103c-1102c(mazEF3)、Rv1991a-1991c(mazEF6)和Rv2801a-2801c(mazEF9)属于经典的TAS,其中mazEF6、mazEF9在营养匮乏环境中启动子增强最显著。本研究结果显示北京基因型比非北京基因型菌株的mazEF3基因转录水平高,两者同为临床致病菌,或许mazEF3高表达赋予了北京基因型菌株抗压力强的特点,在入侵宿主时可以抵抗机体的不利环境而生存,这也许是造成其易流行、持留生存及出现耐药菌性的原因之一。当MTB处于不利环境时或入侵宿主遭遇免疫系统攻击时,接受细胞外死亡因子的刺激,细菌的TAS被激活,其中毒素性质稳定,而抗毒素性质不稳定易被蛋白水解酶水解,造成毒素累积,而北京基因型菌株毒素基因均高表达,可产生更多的毒素,可以选择性的使一些细菌死亡,从而为剩余的细菌保留能量和营养物质,此为利他性死亡,以保证群体稳态和适应不利环境。
作为新疆主要流行株北京基因型菌株mazE9低表达,推测为是内在特定分子表达调节的结果,尤其是mazE9低表达和mazF9高表达的协同作用,能使更多的毒素累积进行利他性程序性死亡,从而提高自身对不良环境的耐受性,也许这也是成为流行株的原因之一。
研究证实大量诱导表达MazF可使细胞处于停滞状态,随后大剂量诱导表达MazE可中和MazF的毒性,使细胞恢复活性。这一停滞状体与结核分枝杆菌的休眠状态很相似。当游离的MazF在细胞内时间或累计量超过某一界点时,就会导致细胞的死亡。我们推测毒素蛋白累积量不同可以使细胞表现出不同的状态,当少量累计时提高细胞对不利生存环境的耐受,当环境进一步恶化,毒素持续累积,促使细胞进入休眠状态,当毒素作用时间或者累积量达到某一值时细胞死亡。实验中使用Western Blot技术检测蛋白质水平的表达时,结果显示,北京基因型菌株相对于非北京基因型菌株,MazF9蛋白高表达,与mazF9基因mRNA表达趋势一致。在转录和翻译水平上,mazF9的高表达也许是北京基因型菌株成为流行株、广泛传播的原因之一。
综上所述,作者初步探索了北京及非北京基因型结核分枝杆菌的生长曲线、mazEF3,6,9系统表达有无差异以及mazEF系统是否与北京基因型菌株流行相关。研究结果表明,北京与非北京基因型结核分枝杆菌的生长曲线、mazEF3,6,9系统表达存在差异,mazEF系统与北京基因型菌株流行相关。MTB的染色体上存在着大量的TAS系统,mazEF3,6,9系统只是其中之一,mazEF3,6,9系统在MTB中究竟如何发挥作用以及与其他TAS系统、调节因素的互作用需要进一步研究。
相关新闻推荐
2、不同温度下鲜切草鱼脊肉块中热杀索丝菌的生长规律、动力学模型的拟合及验证