速生集胞藻6803光合放氧速率、生长曲线及基因组分析(五)
在高光条件下,相较于野生型6803,高光6803基因组中编码响应光强变化的转录调控因子RpaB的特有突变基因slr0947的表达水平未检测到变化;PSII反应中心蛋白D1、D2编码基因psbA2、psbA3和psbD的表达分别上调了4.4、14.8、5.3倍,而蛋白D1、D2的再生能力决定了PSII适应光胁迫、光抑制的能力。PET上的重要电子传递组分细胞色素b6f复合体(B6F)由8个亚基组成,连接PSII、PSI间的电子传递通路。高光6803的B6F编码基因petB的表达水平相比野生型6803上调了4.3倍;以上结果可帮助理解高光6803两个光系统处rETR明显高于野生型6803的原因。
蓝藻胞内有2种还原力:NADH和NADPH。不同于异养生物胞内NADH含量更高,蓝藻胞内的NADPH不仅在总量和比例上占据优势,同时也是光反应的产物,参与胞内固碳反应等生化过程。ATP、NADPH都是光合作用的产物,但总体上,NADPH相对于ATP过剩,光强过高时产生的过量NADPH会损伤胞内微环境平衡。铁氧还蛋白(ferredoxin,Fd)是PET的重要组分,依赖携带的铁硫中心发挥功能,也参与线性电子传递(linear electron transport,LET)中NADPH的代谢。在高光培养条件下,相对于野生型6803,高光6803中编码Fd的petF(fdx)的转录下调了93.7%,编码NADH脱氢酶亚基的ndhD2转录上调12.8倍,编码具有NADPH脱氢酶活性的砷抗性蛋白的arsH转录上调55.6倍。此外,编码双向[Ni-Fe]氢化酶复合体的hoxF与hoxU的转录分别下调了76.2%与82.1%。根据以上转录组结果,我们推测高光6803通过加强还原力的利用维持胞内氧化还原平衡,避免高光胁迫发生,保证各生化反应的正常进行,以有效利用高光。
与以上光反应相关基因的差异化表达趋势类似,在高光条件下,相比野生型6803,高光6803中与光合固碳、能量-物质代谢、跨膜转运相关基因的表达水平也有显著变化。如cmpA、cmpB编码ATP依赖的碳酸氢盐转运蛋白复合物组分,cmpA与cmpB的表达分别上调了24.3倍与10.5倍。此外,编码卡尔文循环中核糖5-磷酸异构酶的rpiA表达上调了6.0倍,编码磷酸甘油酸激酶的pgk、编码果糖-1,6-二磷酸醛缩酶的fba表达分别上调21.5倍与7.0倍。因此,我们推测高光6803耐高光、生长快、生物量积累多也得益于碳酸氢盐转运蛋白复合物、固碳反应相关基因的表达上调。
脂类为蓝藻细胞膜、类囊体膜上的主要成分,还参与维持膜结构与功能、细胞储能。在高光条件下,相比野生型6803,高光6803中与脂类合成相关基因fabD、fabG的表达分别上调了4.3倍与12.4倍;编码单葡糖基二酰甘油差向异构酶的mgdE表达上调4.2倍。以上基因的表达水平上调表明,高光6803也可通过提升脂类合成能力来适应高光强环境,维持胞内生化反应的正常进行。
在高光条件下,相比野生型6803,高光6803基因组中与丙酮酸代谢相关的编码磷酸烯醇丙酮酸合酶的ppsA表达下调了86.5%;与糖酵解过程相关的编码果糖磷酸激酶的pfkA(pfkB1)与pfkA(pfkB2)、编码丙酮酸氧化还原酶的nifJ表达分别下调了86.3%、91.3%与95.9%;编码草酸脱羧酶的sll1358表达下调了81.8%。综上所述,在高光培养条件下,相比野生型6803,高光6803中与糖酵解过程、磷酸戊糖途径等分解反应相关的基因表达水平均下调,与光合固碳相关的多个基因的表达水平均上调。以上表明高光6803通过减弱释放能量的糖分解途径,强化高耗能的光合固碳途径来调控胞内能量-物质平衡,以实现对高光的有效利用与快速生长。
在蓝藻中参与氮同化作用的硝酸盐转运蛋白NrtB、NrtC与NrtD负责向蓝藻胞内转运硝酸盐,并且均依赖ATP供给。在高光条件下,相比野生型6803,高光6803基因组上的nrtB、nrtC与nrtD表达水平分别上调了4.6、5.9、6.4倍,编码硝酸盐还原酶的narB表达水平上调了8.7倍;而与氨基酸降解代谢相关的编码氨基甲酸激酶的arcC、编码谷氨酰胺连接酶的glnA和glnN、编码苏氨酸合成酶的thrC转录水平下调了73.0%–87.3%。综上,我们推测在高光条件下,高光6803的氨基酸分解代谢减弱,氮同化过程增强,可以利用光反应吸收的一部分能量,有利于胞内能量平衡的维持,减轻胁迫,帮助细胞实现快速生长。
在蓝藻与外界进行的物质交换过程中,ATP依赖的ABC转运子蛋白复合体是不可或缺的。在高光条件下,相比于野生型6803,高光6803基因组中除了上述表达水平显著上调的硝酸盐、亚硝酸盐和碳酸氢盐转运蛋白编码基因外,差异化表达的ABC转运子组分编码基因的转录多数都上调了至少4倍(4.2–24.3倍)。因此,相比野生型6803,高光6803表现出更旺盛的跨膜物质运输,这有助于其更高效地利用高光、快速生长、抵抗胁迫。
当蓝藻细胞受外界胁迫时,胞内的DNA复制、修复、转录、重组等生化过程会受到不同程度的影响。在高光条件下,相比野生型6803,高光6803基因组中的编码GIY-YIG核酸酶家族蛋白基因sll0441(SGL_RS18010)、编码NgoFVII家族的限制性内切酶基因slr6047(SGL_RS01585)的表达水平分别下调了80.8%与90.7%。编码DNA解旋酶RecQ的基因recQ表达水平相比野生型6803下调了87.2%,已知依赖ATP的DNA解旋酶RecQ参与DNA复制、DNA修复、mRNA合成及RNA降解等过程。这或许提示我们,在面对胁迫时,相比野生型6803,高光6803耐受性更强,故其所受伤害更小,对DNA重组修复、错误RNA降解的需求也更低。
相比野生型6803,在高光条件下,高光6803中特有的突变基因青霉素结合蛋白1B编码基因sll1434(mrcA)的表达下调了79.2%;与细胞运动相关的鞭毛基因pilT2的表达水平下调了80.0%。这提示我们,高光6803可通过改变或调控细胞壁结构来适应环境变化、利用高光实现快速生长。
综上所述,在对获得的转录组数据进行深入分析后,我们发现在高光900μmol/(m2·s)条件下,为高效利用高光,维持胞内能量、物质代谢平衡,实现快速生长,高光6803几乎调动了所有功能途径。例如,在光合作用的光反应阶段,高光6803胞内PSII反应中心蛋白D1、D2的合成能力增强,编码发挥电子传递功能的细胞色素b6f复合体组分的基因表达上调;NADPH和NADH的利用和分解过程也被加快,以促进从光能到化学能ATP的转化;减轻光损伤,提升PSII的光化学活性。此外,在高光条件下,高光6803胞内与固碳反应相关的基因表达显著上调,而糖酵解及磷酸戊糖途径等脱碳反应相关基因的表达则显著下调,由此可知,当面对高光时,高光6803通过加强光合作用暗反应中的固碳过程,加强可大量消耗能量的氮同化功能、脂类合成及跨膜物质运输,减弱糖酵解及磷酸戊糖途径等脱碳反应,从而维持细胞能量-物质代谢过程的总平衡,实现细胞对高光的高效利用与快速生长。
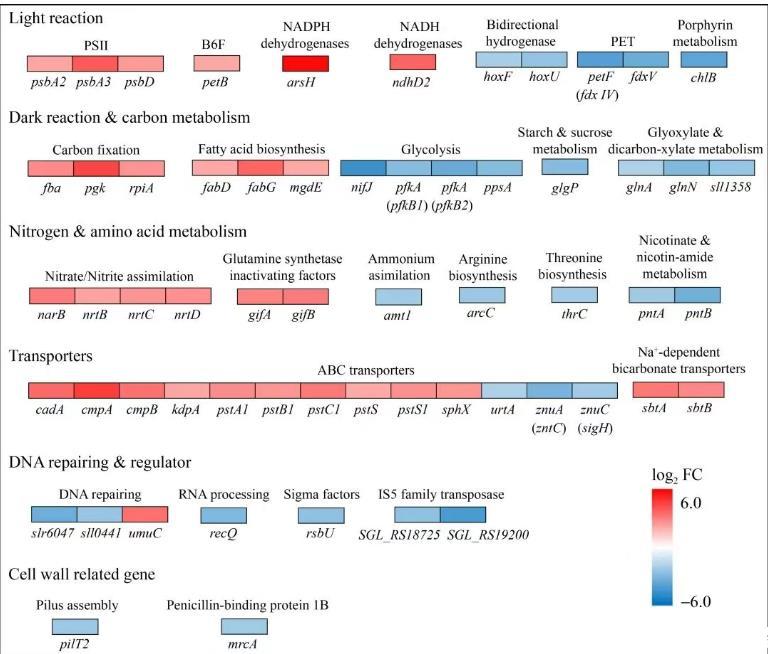
图6在900μmol/(m2·s)高光强下高光6803相对野生型6803的基因差异化表达以蓝色表示基因表达下调,以红色表示基因表达上调;PET:光合电子传递;FC:基因差异化表达的倍数;log2 FC>1。
相关新闻推荐
2、单一外源微生物的生长繁殖情况研究——诱变菌B38的生长曲线绘制
3、基于ELISA和细菌生长曲线应用的结合定量检测肠炎沙门氏菌(二)